
Hogyan alakultak ki a baktériumok, gombák és növények az ősi esők idején
Az esőképződés biológia történelme


A baktériumok és a gombák már jóval a növények és állatok megjelenése előtt jelen voltak a Földön, és megteremtették az alapot a későbbi fejlődéshez.
Lynn Margulis biológus, aki kulcsfontosságú felfedezést tett az eukarióták eredetéről és a szimbiózis evolúcióban betöltött fontos szerepéről, pályafutásának nagy részét azzal töltötte, hogy azt állította: a baktériumok nem csupán a korai életformák voltak, amelyeket később összetettebb formák váltottak fel. Ők voltak – és ma is azok – az alapréteg, amelyen minden más működik; inkább egy operációs rendszerhez hasonlítanak, mint egy primitív ősökből álló csoporthoz. A baktériumok mindenütt jelen vannak: a talajban, az óceánokban, a növények és állatok szöveteiben, a kontinensek feletti levegőben és a gleccserek jégében egyaránt. Beépültek a korai eukarióták sejtjeibe, és azokból lettek a mitokondriumok, amelyek ma szinte az összes összetett életformának energiát szolgáltatnak. Átszövik a növények gyökereit, bélelik az állatok beleit, és olyan mennyiségben keringnek a légkörben, amelyet csak most kezdünk el mérni.
Margulis szerint ahhoz, hogy megértsük, hogyan alakult ki a Föld légköre és éghajlata, meg kellett értenünk, hogyan fejlődtek együtt velük a mikrobák – nem mint egy fizikai bolygó utasai, hanem mint aktív résztvevők, akik alakították annak kémiai összetételét, gázait, éghajlatát és az élet feltételeit. Véleménye szerint a bolygó és mikrobiómája nem voltak egymástól elválasztható dolgok. Milliárd évek óta kölcsönösen alakították egymást.
A levegő, amit belélegzünk, nagyrészt mikrobiális eredetű. A Nagy Oxigenizációs Esemény – azaz a Föld légkörének a fotoszintetikus baktériumok által mintegy 2,4 milliárd évvel ezelőtt végbement átalakulása – tette lehetővé az aerob élet kialakulását, és mindmáig ez az egyetlen legdrámaibb beavatkozás, amelyet bármely életforma bolygószintű méretekben végrehajtott.
A légkör mikrobiális alakítása azonban nem állt meg ennél. A nitrogén a talajban élő denitrifikáló baktériumok révén kerül körforgásba; a levegőben a nitrogén és az oxigén aránya több milliárd éven át tartó mikrobiális nitrogénmegkötés és denitrifikáció eredménye, amelyek anyagcsere-folyamatokban egymással ellentétes irányban zajlanak. A metán, amely oxigénben gazdag légkörben nem maradhatna fenn – mivel reakcióba lép az oxigénnel és egy évtizeden belül eltűnik –, az óceáni üledékekben, a vizes élőhelyeken és az állatok belében élő metánt termelő ősbaktériumok által folyamatosan pótlódik.
A baktériumok a szénciklust a légkör felé és onnan ki irányítják, ezzel geológiai időskálán befolyásolva a Föld hőmérsékletét. Ezen folyamatok mindegyike ugyanazt az alapvető valóságot tükrözi, amelyet Margulis azonosított: a mikrobák nem csupán a Föld kémiai rendszereiben élnek, hanem azokat irányítják is.
A víz a légkör részét is képezi, és itt is a baktériumok és a gombák – amelyekhez később a növények is csatlakoztak – alapvető, de még mindig alulértékelt szerepet játszottak.

A proterozoikum korszakban, nagyjából 2 500–541 millió évvel ezelőtt, valamint az ordovicium korszak elején a szárazföld szabad szemmel nézve szinte teljesen élettelen volt. Bizonyos nedves felületeken mikrobiális szőnyegek tapadtak, de a kontinensek kopárak voltak: szikla, szél és fény.
Amikor eső esett, az többnyire nem mállott ásványi felületekre hullott, rövid ideig tócsákba gyűlt, majd lefolyt. Mivel nem voltak gyökerek vagy biológiai struktúrák, amelyek megakadályozták volna az áramlást, a víz a legközelebbi lejtőn a tenger felé folyt. Nagyon kevés szivárgott be a talajba.
Képzeljünk el egy letömörödött, lecserepesedett, élettelenre művelt szántóföldet - nem sokban különbözik ettől.
Növények nélkül nem volt transzpiráció, vagyis nem létezett olyan mechanizmus, amely a vízgőzt a kontinentális felszínekről visszajuttatta volna a légkörbe. Amikor egy nedvességtelített légtömeg az óceán felől a szárazföld belseje felé halad, minden esőzés alkalmával vizet veszít. Mivel nincs semmi, ami ezt a vizet visszajuttatná a légkörbe, az egymást követő esőzések során az előzőhöz képest egyre kevesebb nedvesség áll rendelkezésre. Ez exponenciális csökkenéshez vezet: a partok közelében erős esőzések vannak, amelyek a légtömeg szárazföld belseje felé haladva gyorsan gyengülnek, mígnem a kontinens belseje szinte semmit sem kap. A szárazföld feletti víz körforgása sekély és egyszerű volt: párolgás az óceánból, szállítás a szárazföldre, felszíni lefolyás, visszatérés a tengerbe.
A geológusok magából a kőzetből rekonstruálták ezt az ősi vízrajzot. Az evaporitok olyan üledékes lerakódások, amelyek akkor keletkeznek, amikor a víz elpárolog, és az oldott sókat hátrahagyja; ezek az ősi vízrajz legértékesebb tanúi közé tartoznak. Amikor egy tó vagy sekély tenger teljesen elpárolog, kémiai nyomot hagy maga után: gipsz, kősó és más ásványok rétegei, amelyek a sótartalom növekedésével meghatározott sorrendben kicsapódnak. Jelenlétük és elhelyezkedésük alapján a geológusok meg tudják állapítani, hol volt állóvíz, és mennyi ideig maradt fenn. A folyók geomorfológiája és az árterületi üledékek tovább egészítik ki a képet. Ezek együttesen felvázolják az ősi világ hidrológiai térképét.
Ezután az élet elkezdett a szárazföldre terjedni, és a baktériumok és gombák egyengették az utat. A mikrobiális szőnyegek után jöttek a gombák hifáikkal: olyan vékony szálakkal, amelyek be tudtak fúrni a sziklák mikroszkopikus repedéseibe. Őket követték a zuzmók, a gombák és a fotoszintetikus algák vagy cianobaktériumok ősi társulásai, amelyek savval oldják fel a sziklát, és olyan felületekhez rögzülnek, amelyeken semmi más nem tud megtelepedni. Ők voltak az útépítők, akik az ásványi felületeket részecskékre zúzták, létrehozva az első durva szubsztrátumot, amelybe mások is gyökeret verhettek.
Az első primitív növények, a mohák és a májmohák, a nedves partvidékeken és a part menti síkságokon jelentek meg. Nem voltak mély gyökereik, és a mai értelemben vett leveleik sem. Szorosan a talajhoz tapadtak, így kiszáradásnak voltak kitéve. De mikorrhiza-kapcsolatokon keresztül olyan gombahálózatokhoz kapcsolódtak, amelyek kiterjesztették hatókörüket az ásványi világba: foszfort gyűjtöttek, a víz halvány rétegeit követték nyomon, és kémiai kitartással oldották fel a kőzeteket. Cserébe a növény a napfényből megkötött szénnel táplálta a gombákat. Az energia lefelé áramlott; a víz és a tápanyagok pedig felfelé. Egy kicsi, törékeny növény így gyakorlatilag egy elosztott organizmussá vált, amely annyira teljesen átszövi a talajt, hogy a növény és a gomba közötti határ elvesztette jelentőségét.
Amit ez a konzorcium lassan, apránként, több millió év alatt felépített, az a talajszivacs volt. Az ép gombahálózatokkal és mikrobiális közösségekkel rendelkező talaj strukturált, porózus, biológiailag aktív anyag, amelynek a vízhez fűződő viszonya alapvetően eltér a csupasz kőzetétől. A rá hulló eső nem folyik le, hanem beszivárog. A pórusokban marad, és napok, hetek alatt a gyökerek rendelkezésére áll. A talajszivacs kialakulása lehetővé tette a csapadék nagyobb mértékű újrahasznosítását, így a víz tovább juthatott a szárazföld belsejébe.

Ennek az átmenetnek a geomorfológiai nyomai a kőzetekben is felismerhetők. A szilur–devon korszakbeli szárazföldi forradalom – azaz az a körülbelül 428–359 millió évvel ezelőtti időszak, amikor a vaszkuláris növények először terjedtek el a part menti síkságokon – üledékes sorozatai azt mutatják, hogy a csupasz földterületek vízelvezetésének fonott, epizodikus csatornahálózata először foltokban, majd egyre szélesebb körben kezdett átadni a helyét a finomabb szemcsés, rendezettebb üledékeknek, amelyek stabilizált partokról és tartós áramlásról tanúskodnak. A kontinentális belterületeken az evaporit-lerakódás eloszlása megváltozott: a peremterületeken kevésbé szélsőségessé vált, ami összhangban áll a víz kissé hosszabb tartózkodási idejével és a belföldi csapadékmennyiség mérsékelt növekedésével. A korai érnövények, amelyek a talajból vettek fel vizet, majd transzpiráció útján bocsátották ki, az óceáni nedvesség hatókörét kissé beljebb terjesztették a szárazföld belsejébe. Ezzel csapadék-visszakerülést hoztak létre.
Dan Ibarra – még a Stanfordon posztdoktorként – modellezte ezt a jelenséget, és bebizonyította, hogy a szárazföldi növények evolúciója és transzpirációs fluxusuk növekedése mérhető mértékben növelte a kontinens belsejében hulló csapadék mennyiségét, ami összhangban áll a geológiai leletekkel [Ibarra 2019].
A hatás azonban korlátozott volt. Mély gyökerek és nagy levélfelületek hiányában ezek a korai növényközösségek csak szerény mennyiségű vizet tudtak felfogni és visszajuttatni a légkörbe.
Kissé lelassították a nedvesség exponenciális csökkenését, és a csapadékot egy kicsit beljebb terelték a szárazföld belsejébe, de nem egészen a kontinens belsejéig.
Az igazi átalakulás a devon korszak végén és a karbon korszak elején, körülbelül 359–299 millió évvel ezelőtt következett be, amikor az első erdők elterjedtek a kontinenseken. Ekkor már a Földön magas, mély gyökerekkel rendelkező fák, sűrű lombkoronák és hatalmas mocsaras ökoszisztémák voltak jelen, és ezekkel együtt a víz körforgásának biológiai szabályozása is fokozódott kontinentális szinten.

A geomorfológiai leletek rögzítik ezt az átmenetet. Az egyenes folyórendszerekről a kanyargós folyórendszerekre való áttérés – amely a növényzetnek köszönhető, az ártereket stabilizáló és az áramlást mérséklő hatás jellegzetes nyoma – ezen időszak során széles körben megfigyelhető számos kontinentális rétegsorban. A kontinentális belterületeken az evaporit-lerakódás a karbon korszak során drámaian visszaesett, ami összhangban áll azzal, hogy az esővíz ekkor már messze behatolt a szárazföld belsejébe, a víz tartózkodási ideje jelentősen meghosszabbodott, és a medencék szorosan összekapcsolódtak a vízelvezető hálózatokkal.
Ez a változás teljesen más rendű volt, mint amit a korai érnövények idéztek elő.
Míg azok a korábbi növényközösségek csupán lassították a nedvesség szárazföld belsejébe történő elpárolgását, az erdők megakadályozták azt.
Egyetlen nagy fa naponta több száz liter vizet tud elpárologtatni több levélrétegen keresztül, visszajuttatva azt a légkörbe, hogy azután a szárazföld belsejében ismét lehulljon. Az erdő ezt több millió fa segítségével egyszerre végzi, a nedvességet a lombkoronától a felhőig, majd az esőig továbbítva egy olyan láncreakcióban, amely az óceán vizét mélyen bejuttathatja a kontinens belsejébe, amely korábban hidrológiai szempontból holtzónának számított.
Az erdők hozzájárulnak az eső kialakulásához. Az erdők azonban nem léteznek elszigetelten. Egy olyan baktériumokból és gombákból álló alaprétegből nőnek ki, amely már sokkal régebb óta létezik, és ez a réteg nem csupán támaszt nyújt az erdőnek, de aktívan részt vesz az eső kialakulásában az erdő mellett.
Az elmúlt fél évszázad során a biológia, az ökológia és a földtudományok területén végbement eszmei konvergencia arra késztette a tudósok egy részét, hogy az életet ne annyira egyedekből álló egésznek tekintsék, hanem inkább összetett rendszereknek, olyan réteges rendszereknek, amelyekben a mikrobák és a gombák nem mellékszereplők, hanem alapvető fontosságú elemek.
A növények és az állatok nem állnak elkülönülve ettől a mikrobiális világtól.
Ez a világ alkotja őket, és ez tartja fenn őket.
Lynn Margulis endoszimbiózissal kapcsolatos kutatásai kimutatták, hogy minden növény és állat sejtjei maguk is egykor független baktériumok összeolvadásából jöttek létre. A sejtjeinket energiával ellátó mitokondriumok, valamint a növények fotoszintézisét lehetővé tevő kloroplasztok nem a semmiből jöttek létre, hanem beépültek a sejtekbe. Ha a komplex élet legalapvetőbb egységei már eleve szimbiotikusak, akkor a tiszta, autonóm organizmus fogalma kezd összeomlani.
A talajban, ahol a szárazföldi élet nagy része kezdődik, a gyökerek és szárak látható világa egy sűrű, dinamikus mikrobiális mátrixba ágyazódik.
A baktériumok és gombák lebontják a kőzeteket, újrahasznosítják a tápanyagokat, és kialakítják azt a fizikai környezetet, amelyben a növények növekednek. A növények nem csupán tápanyagokat vonnak ki ebből a világból, de összefonódnak vele. Gyökereiken keresztül szén-dioxidot cserélnek ásványi anyagokra, alakítva a körülöttük lévő mikrobiális közösségeket, miközben ezek a közösségek is meghatározzák, hogy a növény mire férhet hozzá.
Suzanne Simard erdei ökológus kimutatta, hogy a mikorrhiza gombákkal összekapcsolt fák szén-dioxidot és tápanyagokat továbbítanak egymás között, néha ezzel támogatva a fiatalabb vagy stresszhatásnak kitett szomszédjaikat [Simard 1997, 2004].
Így tekintve az erdő nem versengő egyedek gyűjteménye, hanem egy összekapcsolt hálózat, amelynek tagjait a gombás csatornákon átáramló anyagáramok kötik össze. A versengés és a konfliktusok továbbra is fennállnak, de az élet egysége nagyobb és összetettebb, mint amilyennek első pillantásra tűnik.
Scott Gilbert biológus a holobiont fogalmának segítségével járult hozzá e gondolkodásmód rendszerezéséhez: ez egy olyan organizmust jelöl, amely az összes hozzá kapcsolódó mikroorganizmussal együtt egyetlen ökológiai és evolúciós egységként működik. A fa nem csupán fa-szövet.
A fa, a gombák és a mikrobióma együttese, amely a gyakorlatban elválaszthatatlan, még ha elméletileg meg is különböztethető.
Ez a nézőpontváltás fontos, amikor azt kérdezzük, ki hozza létre az esőt. Ha az élet egysége a holobiont, akkor a transzpiráció nem csupán a fa tevékenysége, hanem az egész rendszeré: a növényi szövetek biztosítják a levélfelületet és a vaszkuláris szerkezetet, a mikorrhiza gombák szabályozzák a víz gyökereken keresztüli áramlását és a sztómák nedvességstresszre adott reakcióját, a baktériumok pedig befolyásolják azokat a molekuláris jeleket, amelyek irányítják a pórusok nyitását és záródását.
Mohanned Abdalla talajfizikus és Mutez Ali Ahmed kertészeti kutató megmérték, hogy az arbuszkuláris mikorrhiza gombák hogyan befolyásolják a paradicsom transzpirációs sebességét különböző nedvességviszonyok mellett [Mohanned 2012]. Robert Augé, Heather Toler és Arnold Saxton a Tennessee-i Egyetemen azt találták, hogy ezek a gombák befonódnak a növényi gyökerekbe, és szabályozó hatást gyakorolnak a sztóma vezetőképességére, azaz arra a folyamatra, amelynek során a növény megnyitja levélpórusait a légzés és a transzpiráció érdekében [2015].
A Kaliforniai Egyetem Riverside-i kampuszán dolgozó Shushu Jiang és munkatársai megállapították, hogy a baktériumok fehérje-jelátviteli útvonalakon keresztül befolyásolják a transzpirációt [2013].
Ezek a kutatócsoportok nem azt vizsgálták, hogy ez mit jelent az esőzések szempontjából.
Az általuk azonosított mechanizmus azonban minden szinten működik. Több millió fa esetében a gombák és baktériumok által szabályozott sztómás viselkedés határozza meg a transzpiráció időzítését és mennyiségét. A transzpiráció tehát nem pusztán botanikai folyamat, hanem mikrobiális és gombás is egyben.
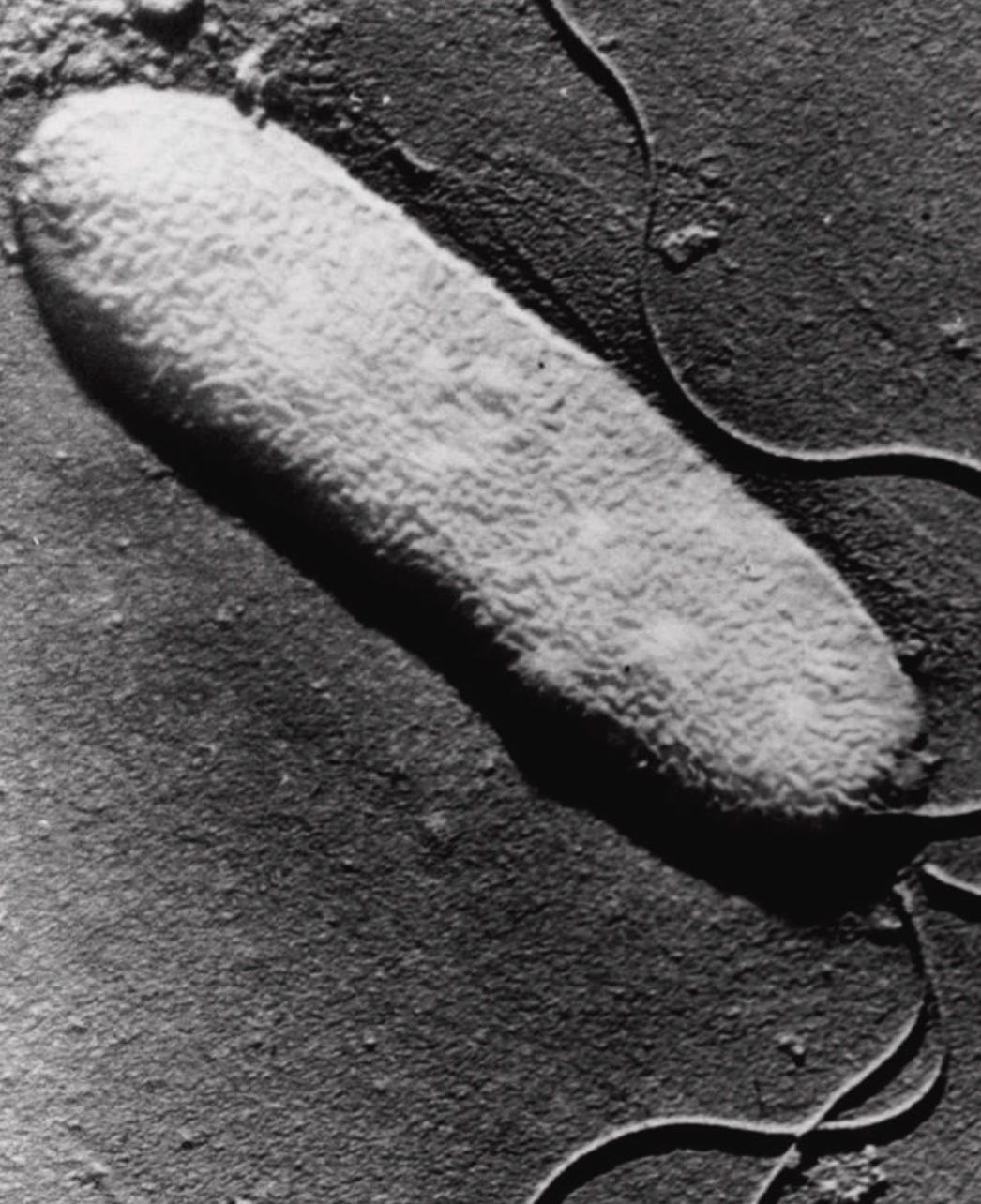
A felhőcseppek nem alakulnak ki pusztán azért, mert a páratartalom elég magas. Ehhez szükség van egy kis részecskére is – amelyet aeroszolnak neveznek –, amelyen a cseppek kialakulhatnak.
A meteorológusok eddig inkább a szervetlen anyagokra összpontosítottak az esőkialakulás szempontjából, de a mikrobák és a gombaspórák is elősegíthetik az eső kialakulását. A baktériumok esőcsapadék-kiváltó képességét először Gabor Vali és Russell Schnell fedezte fel az 1970-es években, majd az 1980-as években újra felfedezte David Sands növénypatológus, aki egy Petri-csészével a kezében lógott ki egy kétfedelű repülőgépből, hogy baktériumokat gyűjtsön a felhőkből.
Egy bizonyos baktérium, a Pseudomonas syringae felszínén olyan fehérjeminta volt, amely segítette a vízmolekulák jégszerkezetekké történő átrendeződését.
A baktérium külső membránján található fehérjék fizikai sablonként működnek, amelyek a vízmolekulákat már -2 °C-os hőmérsékleten is kristályos jégrácsba kényszerítik – ez jóval melegebb, mint a -15 °C, amely általában szükséges ahhoz, hogy a szervetlen aerosol, mint a por kiváltsa a fagyást. Az az elképzelés, hogy baktériumok és gombaspórák esőmagvakként működhetnek, eleinte szokatlannak tűnt, de az évtizedek múlásával egyre inkább elfogadottá vált a klímatudományban.
Kim Prather légköri tudós, aki az Egyesült Államok egyes részein felderítő repüléseket végzett, megállapította, hogy az esőmagvakként működő részecskék harmada biológiai eredetű volt. Christian Pöhlker és kollégái az Amazonas távoli területeire utazva azt találták, hogy a bioaeroszolok – azaz a gombaspórákat, baktériumokat és erdei terpéneket tartalmazó szerves anyagok, amelyek esőt indítanak el – még gyakoribbak voltak az érintetlen erdő levegőjében, ahol a biológiai és szervetlen csírázóanyagok aránya jóval magasabb volt, mint a szennyezett vagy leromlott régiókban [2012].
Az őskorban, az ipari szennyezés és a nagyméretű erdőirtás előtt, várhatóan a baktériumok és a gombaspórák lényegesen nagyobb szerepet játszottak az eső kialakulásában, mint manapság. [További információkért lásd a bioaeroszolok tudományáról és felfedezéséről szóló cikkeimet I. rész, II. rész, és III. rész]
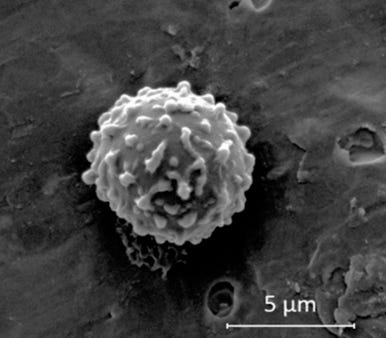
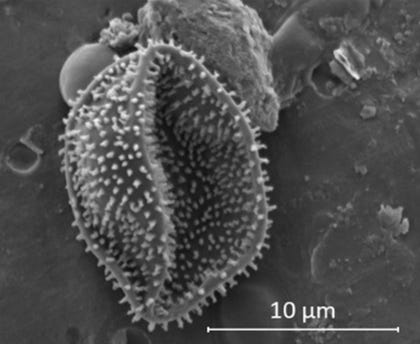
Az erdők maguk is apró szerves részecskéket bocsátanak ki. A tűlevelűek, a lombos fák és sok más növény illékony szerves vegyületeket, terpéneket bocsát ki – ezek azok a molekulák, amelyek a fenyőerdők gyantás illatáért felelősek –, amelyek a légkörben az ózonnal és a hidroxil-gyökökkel reagálva másodlagos szerves aeroszolokat képeznek, amelyek méretüknél fogva felhőkondenzációs magként működhetnek.
Az erdők folyamatosan termelnek olyan részecskéket, amelyek körül felhőcseppek alakulnak ki, és ezzel kémiailag megváltoztatják a fölöttük lévő légkört, így növelve az esőzés valószínűségét éppen azon a területen, ahol az aeroszolok keletkeztek.
Cindy Morris mikrobiológus a baktériumok bioaeroszol-képességeinek evolúcióját a mélyidő perspektívájából vizsgálta [Morris 2012]. A virágos növények és a hatalmas erdők megjelenése olyan hőáramlatokat hozott létre, amelyek a baktériumokat magasra emelték a légkörbe, míg a Pangea szétesése új partvonalakat és hűvös, nedves, vegyes fázisú felhőket hozott létre, amelyekre a baktériumoknak szükségük van a csapadék kiváltásához. Ebben a környezetben a szelekció a jégkristályosító képességet részesítette előnyben a nagy távolságú terjedés megoldásaként: azok a baktériumok, amelyek felhőket tudtak bevetni és eső formájában saját leereszkedésüket kiváltani, nagyobb valószínűséggel jutottak el friss növényi gazdákhoz. Miután a földre kerültek, az általuk elősegített esőből származó nedvesség elősegíti a populáció robbanásszerű növekedését, gyakran ezerrel növelve a baktériumok számát 48 óra alatt, míg a jégkristályosodásból származó fagy felhasítja a növényi sejteket, felszabadítva a tápanyagokat. Ezen visszacsatolási kör révén ezek a mikrobák a szárazföldi növényekkel együtt fejlődtek, hogy elősegítsék mindkettőjük számára szükséges nedves, mérsékelt éghajlati viszonyok kialakulását.
Hasonló logika vonatkozik a gombaspórákra is. Azok a spóra-vonalak, amelyek hatékony jégkristályképzőként működnek, előnyben részesülnének a szelekció során, mivel az eső kiváltása növeli annak esélyét, hogy nedves, fogékony talajra hulljanak, ahol a csírázás lehetséges. A spóratípusok sokfélesége egy adott légtömegben nem a rendszer zaját jelenti, hanem funkcionális változatosságot, ahol az esőindító szakosodott spórák egy feladatot látnak el, míg más vonalak más feladatokat. Egyesek a hosszú légköri utazásra vannak optimalizálva; mások a leszállás utáni gyors csírázásra; megint mások pedig a megzavart talajban a mikorrhiza-hálózatok gyors kialakítására. Az ökoszisztéma mind az esőindítókból, mind a letelepedőkből profitál.
Az a kérdés, hogy egy ilyen rendszer hogyan alakult ki, arra utal, hogy a szelekció egyszerre több szinten is működik.
Az egyes szervezetek szintjén egy olyan baktérium, amely képes jégkristályokat képző magként működni a felhőkben, nagyobb valószínűséggel kerül le a csapadékkal egy nedves növényi felületre, ahol szaporodni tud. Egy olyan gombaspóra, amely hatékonyan elősegíti az eső kialakulását, nagyobb valószínűséggel landol fogékony, nedves talajon, ahol csírázás lehetséges. Egy olyan mikorrhiza-hálózat, amely a helyi páratartalmat fenntartó módon szabályozza a transzpirációt, nagyobb valószínűséggel marad fenn egy élő, termékeny erdőben, mint egy olyan, amely hagyja, hogy gazdaszervezete kiszáradjon. Minden egyes organizmust olyan tulajdonságai alapján választ ki a természetes szelekció, amelyek véletlenül hozzájárulnak a nedvességkörforgáshoz. Egyetlen egyednek sem kell szándékosan törekednie a nagyobb hatásra. Cindy Morris pontosan ezt az érvet hozta fel a jégképző baktériumok esetében: esőcsapadékot előidéző képességük nem véletlen, hanem egy terjedési stratégia, amelyet a szelekció több száz millió év alatt finomított ki.
Az egyéni szelekció azonban önmagában nem ad teljes magyarázatot a megfigyelt integrációra. A csoportszelekciót – amelyet George Williams 1960-as évekbeli kritikái után nagyrészt elvetettek – az elmúlt évtizedekben jelentősen rehabilitálták az evolúciós kutató David Sloan Wilson többszintű szelekciós keretrendszerének, valamint a főbb evolúciós átmenetek vizsgálatának köszönhetően. Különösen az ökológiai rendszerekben egyre inkább elismerik, hogy a közösség olyan valódi egység, amelyre a szelekció hatással lehet.
Egy olyan erdei talajközösség, amelyben a mikroorganizmusok, gombák és növények közötti kapcsolatok szorosan összefonódnak – amelynek mikorrhiza-hálózata hatékonyan szabályozza a transzpirációt, baktériumpopulációi aktívan elősegítik az eső kialakulását, gombaspórái és terpén-aeroszoljai pedig előkészítik a fölötte lévő légkört –, sikeresebben fogja megőrizni és új területeket benépesíteni, mint egy ilyen integrációval nem rendelkező közösség.
Több nedvességet vonz majd a szárazföld belsejébe, a száraz időszakokban is fenntartja a termelékenységet, a zavarások után gyorsabban helyreáll, és határait tovább tolja a korábban száraz kontinentális belterületek felé. Az egyéni szintű és a közösségi szintű szelekció ebben az esetben nem egymással versengő magyarázatok, hanem kölcsönösen erősítik egymást.
Az egyes baktériumokat, spórákat és gombahálózatokat olyan tulajdonságok alapján választja ki a természet, amelyek előnyösek a közösség vízkörforgása szempontjából, és azok a közösségek, amelyek több ilyen tulajdonsággal rendelkeznek, felülmúlják és kiszorítják azokat, amelyek nem rendelkeznek velük.
A szárazföldön hulló eső hosszú története arról szól, hogy mit tettek lehetővé a baktériumok és a gombák: hogyan morzsolták szét a kőzetet és alakították ki a talajt, hogyan befolyásolták a transzpirációt és hogyan készítették elő a felhőket, valamint hogy hogyan fejlődtek együtt a növényekkel több százmillió éven át olyan közösségekké, amelyek képesek az esőt mélyen a kontinens belsejébe juttatni, ahová egyébként szinte egyáltalán nem jutna eső.
Ha többet szeretnél olvasni erről a témáról, olvasd el a „A Föld mikrobiómája szabályozza-e az éghajlatunkat?” című cikkemet
Hivatkozások
Abdalla, Mohanned, and Mutez Ali Ahmed. “Arbuscular mycorrhiza symbiosis enhances water status and soil-plant hydraulic conductance under drought.” Frontiers in Plant Science 12 (2021): 722954.
Augé, Robert M., Heather D. Toler, and Arnold M. Saxton. “Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: a meta-analysis.” Mycorrhiza 25, no. 1 (2015): 13-24.
Ibarra, Daniel E., Jeremy K. Caves Rugenstein, Aviv Bachan, Andrés Baresch, Kimberly V. Lau, Dana L. Thomas, Jung-Eun Lee, C. Kevin Boyce, and C. Page Chamberlain. “Modeling the consequences of land plant evolution on silicate weathering.” American Journal of Science 319, no. 1 (2019): 1-43.
Jiang, Shushu, Jian Yao, Ka-Wai Ma, Huanbin Zhou, Jikui Song, Sheng Yang He, and Wenbo Ma. “Bacterial effector activates jasmonate signaling by directly targeting JAZ transcriptional repressors.” PLoS pathogens 9, no. 10 (2013): e1003715.
Morris, Cindy E., Franz Conen, J. Alex Huffman, Vaughan Phillips, Ulrich Pöschl, and David C. Sands. “Bioprecipitation: a feedback cycle linking Earth history, ecosystem dynamics and land use through biological ice nucleators in the atmosphere.” Global change biology 20, no. 2 (2014): 341-351.
Pöhlker, Christopher, Kenia T. Wiedemann, Bärbel Sinha, Manabu Shiraiwa, Sachin S. Gunthe, Mackenzie Smith, Hang Su et al. “Biogenic potassium salt particles as seeds for secondary organic aerosol in the Amazon.” Science 337, no. 6098 (2012): 1075-1078 https://www.science.org/doi/abs/10.1126/science.1223264
DC Sands, VE Langhans, AL Scharen, G de Smet, The association between bacteria and rain and possible resultant meteorological implications. J Hungarian Meteorol Serv 86, 148–152 (1982)
Schnell, Russell C., and Gabor Vali. “Looking back: An account of how ice nucleation by bacteria was discovered (1963 to about mid-1980s). Part II: Broadening the scope.” Bulletin of the American Meteorological Society 105, no. 6 (2024): E1004-E1014. https://journals.ametsoc.org/view/journals/bams/105/6/BAMS-D-23-0115.1.xml
Simard, S., Perry, D., Jones, M. et al. https://www.nature.com/articles/41557 Nature 388, 579–582 (1997). https://doi.org/10.1038/41557
Suzanne W Simard and Daniel M Durall. 2004. Mycorrhizal networks: a review of their extent, function, and importance. Canadian Journal of Botany. 82(8): 1140-1165. https://doi.org/10.1139/b04-116
Vali, Gabor, and Russell C. Schnell. “Looking back: An account of how ice nucleation by bacteria was discovered (1963 to about mid-1980s). Part I: The basics.” Bulletin of the American Meteorological Society 105, no. 4 (2024): E778-E788. https://journals.ametsoc.org/view/journals/bams/105/4/BAMS-D-23-0114.1.xml
 | A guest post by
|
Már vártam a posztokat az 50 dodóért, még mindig megéri 😁